
Vol. 36, n.º 2, 2003
|
REVISTA
ESPAÑOLA DE
Vol. 36, n.º 2, 2003 |
Julio Escalona-Zapata, Ana María Calvo de Mora Álvarez, M.ª Dolores Díez Nau, César Lacruz Pelea
Servicio de Anatomía Patológica. Hospital General Universitario Gregorio Marañón. Madrid.
RESUMEN
El gliosarcoma es un tumor cerebral poco frecuente que viene definido por una proliferación bifásica en la que toman parte: a) una fase gliomatosa, habitualmente un glioblastoma, con expresión de GFAP y negatividad para la Vimentina; b) una fase conjuntiva con expresión de Vimentina y negatividad a la GFAP.
Por su parte, el fibrosarcoma cerebral primitivo es un tumor muy infrecuente, que se define por una proliferación de células fusiformes, capaces de expresar Vimentina, incapaces de expresar GFAP y que producen abundantes fibras reticulares y, ocasionalmente, colágenas.
En nuestro medio hemos estudiado dos casos de gliosarcoma por medio del cultivo de tejidos y por medio del cultivo orgánico.
En ambos casos, el cultivo de tejidos demostró la población dual, en forma de colonias celulares diferentes, aun dentro del mismo cultivo. Por su parte, el cultivo orgánico demostró el crecimiento doble en las primeras fases del crecimiento, en tanto que el cabo del tiempo, la fase conjuntiva va predominando sobre la glial, hasta que esta última desaparece.
En consecuencia, se plantea la posibilidad de que el fibrosarcoma primitivo cerebral represente la fase final de un gliosarcoma en el que el componente conjuntivo ha hecho desaparecer el componente glial inicial.
Palabras clave: Gliosarcoma, fibrosarcoma, cultivo de tejidos, cultivo orgánico, overgrowth.
ABSTRACT
Gliosarcoma is an infrequent brain tumor defined by a biphasic proliferation involving: A) a gliomatous stage, usually a glioblastoma expressing GFAP and Vimentin-negative; B) a connective stage with Vimentin expression and GFAP-negative.
On the other hand, primitive brain fibrosarcoma is a very infrequent tumor defined by a proliferation of spindle cells expressing Vimentin but not GFAP, producing abundant reticulin and occasional collagen fibers.
Two gliosarcomas have been studied both by tissue and organ culture. In the two cases, tissue culture showed a dual cell population structured in separate proliferation cell groups, even within the same culture. Organ culture showed a double growth in early stages, whereas later on, the connective stage prevailed over the glial stage, until the latter disappeared.
In conclusion, primitive brain fibrosarcoma might be the final stage of a gliosarcoma in which the connective phase has led to the disappearance of the initial glial component.
Key words: Gliosarcoma, fibrosarcoma, tissue culture, organ culture, overgrowth.
INTRODUCCIÓN
El fibrosarcoma primitivo encefálico es una neoplasia muy infrecuente que no sobrepasa el 0,7 al 1,9 % del total de procesos expansivos intracraneales (1,2) Si bien el número real debe ser menor, ya que algunas casuísticas incluyen todos los tumores sarcomatosos, incluidos los extraaxiales. Morfológicamente, es un tumor monofásico que se define por una proliferación de elementos fusiformes, con variable grado de atipia según los casos y con una tasa mitósica elevada, que tienden a asociarse en fascículos variablemente entrecruzados. Los vasos son capilares de fina pared, ocasionalmente de luz amplia, carentes de los habituales fenómenos proliferativos de los tumores malignos gliales. Característicamente, alrededor de cada célula existe un abundante intersticio colágeno, demostrable tanto con métodos tricrómicos como para la reticulina (3) (fig. 1 a y b).
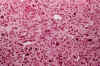
Fig. 1a: Proliferación de
elementos fusiformes con cierto grado de polimorfia y crecimiento irregularmente
fascicular. Los vasos son muy imperfectos a modo de hendiduras tapizadas por
endotelio no hiperplásico. H-E x200.
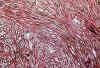
Fig. 1b: Abundante
intersticio reticulínico que hace destacar la disposición fascicular.
Reticulina x100.
Por su parte, el gliosarcoma es un tumor también infrecuente con una incidencia alrededor del 2% (4), con un crecimiento bifásico con áreas gliales y sarcomatosas en proporción variable según los casos (5-8). La fase glial suele ser un glioblastoma, con elevada densidad celular, frecuentes atipias celulares, elevada tasa mitósica y necrosis de diversa amplitud. Tal y como corresponde a este tipo de tumores, estas áreas expresan GFAP en sus células más diferenciadas, carecen de intersticio colágeno y presentan intensos fenómenos de hiperplasia endotelial y adventicial en los vasos. La fase glial no siempre es un glioblastoma, sino que en un pequeño número de casos, es un astrocitoma de distinto grado de malignidad o, incluso, un oligodendroglioma o un ependimoma (9-11).
Las áreas sarcomatosas se disponen mezcladas con las áreas gliales y suelen estar en íntima relación con las zonas de hiperplasia vascular. En ellas, las células son fusiformes o moderadamente estrelladas, con un núcleo central ovalado y yacen en un intersticio rico en fibras conjuntivas. La vascularización es abundante, con vasos de endotelio bien visible, alrededor del cual, las células proliferantes se disponen formando rodetes. Casi siempre hay un buen límite entre las dos fases. Característicamente, las células de estas áreas no expresan GFAP y sí, por el contrario, Vimentina.
Mientras que el origen de las áreas gliales presenta la misma problemática del resto de los gliomas, el origen de las áreas sarcomatosas ha sido objeto de múltiples interpretaciones. La teoría más clásica atribuye su origen a la transformación sarcomatosa de los vasos hiperplásicos de un previo glioblastoma y, por lo tanto, debe admitirse una estirpe originaría endotelial o conjuntiva, según procedan del propio endotelio o de la adventicia vascular, pero estudios subsiguientes han abogado por un origen a partir de pericitos, histiocitos, células musculares lisas y células musculares estriadas. Los estudios de biología molecular han llevado a considerar la fase sarcomatosa como de origen glial, de modo semejante a lo que sucede en las dos variantes desmoplásticas infantiles, el ganglioneuroma y el astrocitoma.
Sin embargo, no hay estudios encaminados dilucidar una posible relación entre el gliosarcoma y el fibrosarcoma intracraneales.
En nuestro medio hemos empleado la técnica del cultivo orgánico en dos casos que se exponen a continuación.
MATERIAL Y MÉTODOS
Para el presente trabajo se han estudiado dos casos de gliosarcoma procedentes del Servicio de Anatomía Patológica del Hospital General Universitario. Gregorio Marañón (Madrid), cuyas características clínicas se resumen a continuación:
Caso 1. Varón de 60 años que consulta por cefaleas y síndrome de hipertensión intracraneal. El estudio neurorradiológico demuestra un tumor temporal derecho que se interviene, con el diagnóstico de gliosarcoma.
Caso 2. Hembra de 64 años que consulta por cefaleas y síndrome de hipertensión intracraneal. El estudio neurorradiológico demuestra un tumor temporooccipital izquierdo que se interviene, con el diagnóstico de gliosarcoma.
Ambos casos se estudiaron con métodos convencionales- H-E; Van Gieson para fibras colágenas; Rio-Hortega para reticulina, e inmunohistoquímicos, GFAP para células astrocitarias y Vimentina para células conjuntivas. Parte de ambos tumores se extendieron para practicar estudios citológicos, los cuales se estudiaron con H-E.
Igualmente, parte de ambos tumores se explantaron para estudio con cultivo de tejidos por medio de la técnica de Kersting (12), modificada por Escalona Zapata y Diez Nau (13).
Por último, ambos casos fueron estudiados en sistemas de cultivo orgánico con arreglo a la siguiente pauta (14):
Fragmentos de 0,3-0,6 cm se explantan en condiciones de esterilidad en placas de Petri de 5 cm de diámetro, colocando cada uno sobre un cuadrado de esponja de fibrina de 0,8 cm de lado, con un total de 3-4 explantos en cada placa. Este material se cubre con medio nutritivo hasta dejar sólo una mínima parte del explanto por encima de su superficie. El medio nutritivo se compone de CMRL 1066 suplementado con un 5% de suero de ternera, bicarbonato sódico, L-glutamina, insulina, hidrocortisona y antibióticos. Este medio caduca a los 7 días de modo que hay que renovarlo cada semana.
Las placas de Petri se colocan en una cámara semiestanca, que yace sobre un agitador de vaivén, a razón de una oscilación cada 4 minutos, de forma que los explantos están en contacto, alternativamente, con el medio de cultivo y con el medio gaseoso.
Todo este sistema se incuba a 37º C en una estufa con medio gaseoso conteniendo un 5% de CO2.
Durante el primer mes de cultivo se separa un explanto cada 5 días y a partir del segundo mes se cada 10 días. Se fijan en alcohol y se incluyen en parafina, practicándose las coloraciones siguientes: H-E; GFAP, Vimentina.
RESULTADOS
Desde el punto de vista histológico, las imágenes corresponden a sendos tumores bifásicos, con una fase glioblastomatosa clásica, con células fusiformes o estrelladas dispuestas al azar con una atipia de grado medio y una tasa mitósica también intermedia. Entre ellas hay una sustancia intercelular desprovista de componente colágeno, apareciendo por el contrario algunos vasos de pared hiperplásica.
Estas áreas alternan con otras en las que las células son fusiformes y se disponen más apretadamente. Entre ellas hay un abundantísimo intersticio colágeno demostrable por medio de técnicas tricrómicas, P.A.S. y reticulina así como una vascularización de capilares de endotelio ligeramente tumefacto, alrededor del cual se densifican las fibras y las células. La actividad mitósica es aquí menor que en las áreas gliales (fig. 2 a y b).
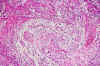
Fig. 2a: Crecimiento mixto
con zonas gliales más claras, de elementos polimorfos carentes de intersticio
fibrilar, que alternan con áreas sarcomatosas con abundante intersticio P.A.S.
positivo. P.A.S.-Hematoxilina x100.

Fig. 2b: Las técnicas para
la reticulina demuestran las dos fases, de las que la glial carece de
intersticio fibrilar, mientras que la mesenquimal posee numerosas fibras
conjuntivas. Reticulina x100.
El estudio citológico de ambos casos muestra repetitivamente la dualidad del proceso, observándose grupos celulares independientes, unos de morfología sarcomatosa y otros de morfología gliomatosa, tal y como sucede en este tipo de tumores (15) (fig. 3 a y b).

Fig. 3a: En la citología,
las áreas gliales muestran una imagen típica de glioblastoma, con elementos
polimorfos, fusiformes o estrellados con núcleos variables e hipercrómicos.
H-E x400.
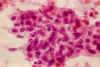
Fig.3b: Las áreas
sarcomatosas muestran una imagen más compacta con cúmulos de células
fusiformes, discretamente polimorfos, sin la asociación reticular propia de los
gliomas. H-E. x400.
En los dos casos estudiados por medio del cultivo de tejidos, las imágenes han sido superponibles a las ya descritas (16-18), observándose explantos distintos dependiendo de la procedencia de las áreas explantadas. En las áreas de procedencia glial, el crecimiento ha sido típicamente astrocítico, con una emigración inicial radial que se transformó rápidamente en reticular. En esta fase de estabilización, las células emigradas eran estrelladas con cierta variabilidad en el tamaño celular, existiendo un cierto número de elementos de gran tamaño y múltiples expansiones que contrastaban con células más pequeñas, a veces bipolares, aun en periodos iniciales de su desarrollo (fig. 4).
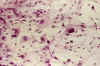
Fig. 4: Cultivo de tejidos a
los 24 días de explantación. Las zonas neuróglicas poseen células
estrelladas muy polimorfas con típica disposición reticular, tal y como es
propio de los glioblastomas. H-E x200.
Por el contrario, las zonas procedentes de las áreas sarcomatosas, presentaron un crecimiento de células bipolares, dispuestas radialmente al principio, para ir formando progresivamente un tapiz más denso, persistiendo la morfología bipolar. Entre estas células, no es raro encontrar algunos elementos aislados, de gran tamaño, con un núcleo central redondo, alrededor del cual se acumulan las organelas y que posee un citoplasma amplio a modo de sábana, que dan a la imagen un gran parecido con las imágenes tipo m de Morley propias de los meningiomas y hemangioblastomas (19,20) (fig. 5).
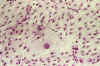
Fig. 5: Cultivo de tejidos a
los 21 días de explantación. Las zonas conjuntivas están formadas por
células más pequeñas, bipolares, dispuestas en tapiz. Ocasionalmente,
aparecen células más grandes, de núcleo central y citoplasma abundante y
plano. La imagen es similar a la de los meningiomas y hemangioblastomas. H-E
x100.
El estudio por medio del cultivo orgánico proporcionó los resultados más llamativos. En los primeros momentos de la vida de los explantos se observaron dos fases simultáneas. En una de ellas, las células eran ricas en citoplasma con un núcleo central o discretamente lateral izado y dispuestas apretadamente, sin apenas sustancia intercelular. Estas áreas expresaron GFAP en su citoplasma y se interpretaron como el componente glial del tumor. Junto a estas zonas, existían otras con células fusiformes de citoplasma más escaso y núcleo más claro. La asociación celular era más fasciculada y no existía expresión de GFAP y expresando Vimentina, por lo que fueron consideradas como la fase mesenquimal del tumor (fig. 6 y 7).

Fig. 6: Cultivo orgánico a
los 20 días. Los explantos muestran una alternancia de áreas gliales, con
expresión de GFAP y áreas negativas, correspondientes a las zonas
sarcomatosas. GFAP x100.

Fig. 7: Cultivo orgánico a
los 50 días. Al cabo del tiempo, las zonas sarcomatosas van predominando,
quedando únicamente algunos pequeños islotes de células con expresión de
GFAP. GFAP x100
Al cabo de varias semanas de cultivo, las células GFAP positivas fueron disminuyendo, quedando reducidas al final, a mínimos islotes inmersos en una gran masa de células negativas para la GFAP y con intensa expresión de Vimentina (fig. 8 y 9).
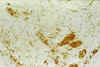
Fig. 8: Cultivo orgánico a
los 20 días. La expresión de Vimentina se reduce, al principio, a algunos
grupos celulares. Vimentina x100.
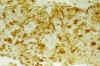
Fig. 9: Cultivo orgánico a
los 45 días. Al cabo del tiempo, las áreas con expresión de Vimentina han
predominado claramente sobre las escasas áreas gliales. Vimentina x100.
DISCUSIÓN
Las investigaciones acerca del gliosarcoma se han centrado básicamente en la naturaleza del componente mesenquimal, la cual ha sido interpretada en diversos sentidos, que pueden resumirse del siguiente modo:
1. La teoría más clásica considera un origen a expensas de los habituales vasos hiperplásicos del glioblastoma, con la siguiente secuencia: vasos proliferantes, hiperplasia máxima endotelial y adventicial, transformación sarcomatosa.
Aunque, en principio se consideró a la célula endotelial como responsable del proceso, la participación de pericitos, células musculares lisas y estriadas e histiocitos (21-23) amplió las posibilidades a todos los componentes de la pared vascular.
2. Un segundo punto de vista defiende la capacidad oncogénica del tejido neoplásico glial sobre los tejidos vecinos no tumorales, de modo que el glioblastoma induciría la transformación sarcomatosa del tejido conjuntivo adyacente (24). En sentido contrario, un sarcoma primitivo podría inducir el crecimiento tumoral glial, dando lugar a un sarcoglioma (25).
3. Más recientemente, las aportaciones de las modernas técnicas han demostrado características semejantes en ambas fases, como las mutaciones en el codon 274 del gen PTEN. Ello unido a la consideración del origen glial del componente mesenquimal en tumores complejos como los ganglioneuromas y astrocitomas infantiles desmoplásicos, del xantoastrocitoma polimorfo y del propio gliofibroma, permitirían extrapolar este concepto monoclonal al gliosarcoma (26).
De otro lado, aunque la proporción entre las áreas gliales y mesenquimales es variable, no existen datos concretos acerca de la evolución dominante en uno u otro sentido. En nuestro estudio, los métodos clásicos, la propia citología y el cultivo de tejidos no han aportado datos nuevos y se han limitado a confirmar la coexistencia de ambos tejidos en el mismo tumor. Solamente el cultivo orgánico ha permitido realizar un estudio evolutivo de la citada relación entre ambos tejidos. En nuestros dos casos, la evolución «in vitro» ha sido la misma con un progresivo predominio del tejido sarcomatoso y subsiguiente disminución de la fase glial, reducida al final a pequeños islotes. Este predominio sólo puede explicarse en base al fenómeno conocido como «overgrowth» tal y como ha sido invocado en otras áreas de la patología (27).
El cultivo orgánico es el único método capaz de proporcionar rediferenciación lo que permite descartar algunos aspectos controvertidos. La rediferenciación excluye que la expresión de Vimentina sea consecuencia de un proceso de desdiferenciación del área glial tal y como sucede en el material «in vivo» (28). Nuestros propios trabajos sobre la rediferenciación del glioblastoma (14,29) han demostrado, al contrario, el aumento de expresión de GFAP a lo largo de la vida del cultivo.
Aplicando todo lo anterior a la naturaleza del fibrosarcoma primitivo cerebral resaltan algunos hechos significativos:
1. Tanto la edad de presentación como la localización del fibrosarcoma y del gliosarcoma son las mismas;
2. Dentro de los gliosarcomas, la proporción entre uno y otro tejido es variable.
3. La morfología de la fase sarcomatosa del gliosarcoma y la del fibrosarcoma cerebral puro son semejantes.
4. Los estudios recientes de biología molecular apoyan el origen monoclonal de ambos tejidos.
Consecuentemente, en base a nuestro estudio, creemos justificado considerar la posibilidad de que el fibrosarcoma primitivo cerebral sea la fase final de la evolución de un previo gliosarcoma en el que la fase sarcomatosa ha destruido la fase glial en virtud del mecanismo conocido como «overgrowth».
BIBLIOGRAFÍA
Cushing H, editor. Intrakranielle Tumoren. Berlín: Springer; 1935.
Zülch KJ. Pathologie der raumbeengenden intrakraniellen Prozessen En: Olivecrona H, Tönnis W, editores. Handbuch der Neurochirurgie, Vol III. Berlin: Springer; 1956.
Escalona-Zapata J. Estudio histopatólogico de los sarcomas de los hemisferios cerebrales. Trab Inst Cajal Invest Biol 1963; 55: 193-227.
Kleihues P, Burger PC, Scheithauer BW, editores. Histological typing of tumors of the central nervous system. WHO Blue Book. 2.ª Ed. Berlin: Springer; 1993.
Feigin I y Gross SW. Sarcoma arising in glioblastoma of the brain. Am J Path 1955; 31: 633-53.
Rubinstein LJ. The development of contiguous sarcomatous and gliomatous tissue in intracranial tumors. J Path Bact 1956; 71: 441-59.
Feigin I, Allen LB, Lipkin L, Gross SW. The endothelial hiperplasia of the cerebral blood vessels with brain tumors and its sarcomatous transformation. Cancer 1958; 1: 264-77.
Morantz RA, Feigin I, Ransohoff J. Clinical and pathological study of 24 cases of gliosarcoma. J Neurosurg 1976; 45: 398-408.
Feigin I, Ransohoff J, Liebermann A. Sarcoma arising in oligodendrogliomas of the brain. J Neuropath Exp Neurol 1976; 35: 679-84.
Escalona-Zapata J, editor. Atlas de Anatomía Patológica de los Tumores del Sistema Nervioso. Madrid: Ed. Complutense; 1986.
Louis DN, Hedley-White ET, Martuza RL. Sarcomatous proliferation of the vasculature in a subependymoma: a follow-up study of sarcomatous dedifferentiation. Acta Neuropathol 1990; 80: 573-4.
Kersting G, editor. Die Gewebszüchtung menschlicher Hirngeschwülsten. Berlin: Springer; 1961.
Escalona-Zapata J, Diez-Nau MD. Estudio de los astrocitomas y sus derivados por medio del cultivo de tejidos. Rev Esp Patología 1983; 16: 23-45.
Calvo de Mora AM. Comportamiento de tumores gliales en sistemas de cultivo orgánico. Estudio convencional e inmunohistoquímico. Madrid: Tesis Doctoral Universidad Complutense; 2000.
Lacruz Pelea C, Escalona-Zapata J, editores. Diagnóstico citológico de los tumores del sistema nervioso. Correlación citohistológica. Barcelona: César Viguera Ed; 2000.
Gullotta F, Kersting G. Sul gliosarcoma. Richerche in vitro. Rev Pat Clin Sper 1964; 5: 205-22.
Gullotta F. Zur in vitro Diagnostik glios-mesenchymaler Mischgeschwülste. Dtsch ZtschrNervenheilk 1964; 186: 323-35.
Bronzini E, Grossi G. La coltura in vitro nella diagnosi del gliosarcoma Pathol 1974; 66: 465-76.
Morley TP. The morphology of meningioma grown in vitro. J. Neuropath Exp Neurol 1958; 17: 643-55.
Escalona-Zapata J, editor. Tumores del sistema nervioso central. Madrid: Ed Complutense; 1996.
Barnard RO, Bradford R, Scott T, Thomas DG T. Gliomyosarcoma. Report of a case of rhabdomyosarcoma arising in a malignant glioma. Acta Neuropathol 1986; 69: 23-7.
Kochi N, Budka H. Contribution of histiocytic cells to sarcomatous development of gliosarcoma. Acta Neuropathol 1989; 73: 124-30.
Haddad SF, Moore SA, Schelper RL, Goeken J. Vascular smooth muscle hiperplasia underlies the formation of glomeruloid vascular structures of glioblastoma multiforme. J Neuropath Exp Neurol 1992; 51: 488-92.
Rubinstein LJ. Morphological problems of brain tumors with mixed cell population. En: Zülch KJ, Woolf PD, editores. Classification of brain tumors. Viena: Springer; 1964.
Lalitha VS, Rubinstein LJ. Reactive glioma in intracranial sarcoma: a form of mixed sarcoma and glioma (sarcoglioma). Report of eight cases. Cancer 1979, 43: 246-57.
Kleihues P, Cavenee WK, editores. Pathology and genetics of tumors of the nervous system. Lyon: IARC; 2000.
Gullotta F, editor. Das sogennante Medulloblastom. Berlin: Springer; 1967.
Schiffer D, Giordana MT, Mauro A, Germano I, Giaccone G. Immunohistochemical demonstration of vimentin in human cerebral tumors. Acta Neuropathol 1986; 70: 209-19.
Escalona-Zapata J, Diez-Nau MD. The astrocytic nature of glioblastoma demonstrated by tissue culture. Acta Neuropathol 1981; 44: 71-5.
![]()